�l(f��)���r�g�����ٷ���r�I(y��)Փ���g�[��1��
ժ Ҫ�� ժҪ�����������܉������е��y�����ף��Y�xɼľ(Cunninghamialanceolata)���H��Ч�������ھ����Ϸ��t���^(q��)ɼľ�˹����������������ކ��}������Ҫ�F�����x.���Ϸ��t���^(q��)��ͬ���g(2��4��10��15a)ɼľ�˹����µĸ��H�������о�����ͨ�^ƽ����x���Y���ꡢҺ�w�l(f��)
����ժҪ�����������܉������е��y�����ף��Y�xɼľ(Cunninghamialanceolata)���H��Ч�������ھ����Ϸ��t���^(q��)ɼľ�˹����������������ކ��}������Ҫ�F�����x.���Ϸ��t���^(q��)��ͬ���g(2��4��10��15a)ɼľ�˹����µĸ��H�������о�����ͨ�^ƽ����x���Y���ꡢҺ�w�l(f��)�͏ͺY�����16SrDNA �y�Y�x���b����Ч���H���������Æ����،�������ԇ�_����Ч�������(y��u)���B(y��ng)�l��.���Y��������1)15aɼľ���H�����Ľ�o�C���c���ЙC�������քe��3.64×105��2.14×105cfu/g�����@�������������g(p<0.05)���Ҹ����g��������g�����@���(p<0.05);2)�Y�x��25���o�C����20����ЙC������ƽ����Y��Һ�w�l(f��)�͏ͺY��16SrDNA �y���b�����քe�õ�һ�����Ч���@��(p<0.05)�Ľ�o�C���� W1(������238.08μg/mL�����ӗU����(Acinetobactersp.))�ͽ��ЙC���� Y9(������15.04μg/mL�����ײ��Ͼ���(Klebsiellasp.));3)��(y��u)���� W1��������B(y��ng)�l����1.0%(�| �� �� ��)�� ���ǡ�1.50%(�|���֔�)��ĸ�ۡ���ʼpH7.5���bҺ��30mL���� �N ��3%(�w �e �� ��)���� ��40 �棬Y9��������B(y��ng)�l����0.5% �����ǡ�1.25%��ĸ�ۡ���ʼpH6.0���bҺ��20mL���ӷN��9%���ض�28��.�����Y���ɞ�ɼľ���H����������_�l(f��)�����ṩ����֧��.
�����P�I�~��ɼľ;����;�Y�x;�b��;���B(y��ng)�l����(y��u)��;�t���^(q��);�Ї��Ϸ�
��������ֲ�����L�l(f��)�����Y���M�ɺ����������^�̵��P�IԪ��֮һ�����r�I(y��)���a�ϳ�ͨ�^����ʩ���ʁ�M�������Ԫ�ص������ʘO���c�����x���γɲ����������}�M���������̶�����[1]���@�O�������˷��ϵ�����Ч��[2-3].��ˣ���λ�������y�ܑB(t��i)�ײ��� �� �� �D �� �� �� Ч �� �� �� �� ǰ �� �� �� �W ���о����c֮һ[4].
�����҇��Ϸ��օ^(q��)��������Ч�����O��[5]��95%��99%�������y�ܑB(t��i)����[6]�������Ƽs���Ϸ���Ҫ�òĘ�Nɼ ľ(Cunninghamialanceolata)�� �� �� �� �� ���m(x��)���I[7-9].ᘌ�ɼľ�˹��ֽ��I�����R�ĵ����{�ȣ��T���W�ߏ�ʩ��[10]����N����[11]�������ʩ[12]�ȷ������� �� �� �� �� �L ԇ���m ȡ �� �� һ Щ �� Ч���� �� �f �o ����Ч���ͳɱ��ظ���ɼľ�˹�����������Ч��ȱ����r.ᘌ��Ϸ��օ^(q��)ɼľ�ֵص��������ކ��}���� ����Ч����ɼľ��������Ч�ʣ���ɼľ�˹��ֵĿɳ��m(x��)���I������Ҫ�ĬF�����x.
�������S��ֲ��-�������|���ϵ�����Ⱥ�䌦�ھS��ֲ�����L�l(f��)����ֲ�。��������Ҫ����[13].�F���о��J����H�������r������H�ĺ��ģ��ڸ��������������|����l(f��)�]�˘O�������[14-16].���ø��H���������ľ���I�B(y��ng)Ԫ�ص����ՠ�r�����T���Lԇ��ǰ���о��� �t �� ��(mangrove)[17]���� β ��(Eucalyptusgrandis)[18]���R β �� (Pinusmassoniana)[19]���� ��(Liquidambarformosana)[20]���� �� �S ̴ (Dalbergiaodorifera)[21]��ֲ �� �� �H �Y �x �� �� һ �� �� �� �@ �� ����Ч���ĸ��H����[22-23]���@������ɼľ�����y���ؗl���µ��B(y��ng)������Ч���ṩ����˼·.�DŽt���[24]���ɼľ�B�Ԍ��������B(y��ng)��������ͣ��M�����F�鲻ͬ���gɼľ�ĸ��H����Ⱥ������^��.ɼľ���H���غ����S�����g�����Ӷ�����[25]�������gɼľ�˹����µ�����������@�������������g[26]���������� �B(y��ng) �� �� �� �� ��.�� �ˣ����о��Բ�ͬ���gɼľ �˹��� �� �� �� �� ���� ɼ ľ �� �H �� �� �� �� ԇ �� ����ͨ �^ƽ�嶨���c�uƿ�����Y�x����Ч�������b����Ч���� �� � ��.�� �� �� �A �ϣ�ͨ�^�����،���c���� ԇ(y��u)����Ч�������L�l�������ګ@�þ����^�߽������������L���ľ��꣬�����ø��H��������ɼľ�˹������H�oЧ����������Ч�ʵ춨�����A�����̽������ �� �� �� �� �� ɼ ľ �� �� �� �� �� ˥ �� �� �} �Ŀ�����.
����1 �����c����
����1.1 ������
������ԇ������Ʒ�ɼ��ڸ���ʡ��ƽ�н�ꖅ^(q��)Ϫ�|�����ֈ�(118°08′��120°31′E��26°40′��27°20′N)����̎�� �� ɽ �} �� �ȣ��� �� �� �� �� �� �� �� �� �� �L �� �����ğᣬ�ļ����������L�l(f��)�_�������ˮ��1700mm�������l(f��)��1500mm������ش���18�棬ʮ��������ɼľ�� �� �L �l(f��) ��.�� �� ǰ �� �� �� �a �� Ч �R β �� �� �� �֣�2008—2010�����꿳����I��ɼͩ�콻��[27]���� ��ֲ�� �N � �� Ҫ �� �� �� (Pleioblastusamarus)��â ݽ(Dicranopterisdichotoma)���^ �� �� ɏ(Strobilanthescyclus)���S��ľ(Adinandramillettii)��.
����1.1.1 ������Ʒ�ɼ����������|�y��
����������xȡ�������g(2��4��10��15a)ɼľ�˹��֣���ȡ3�K���д��� �� ��20m×20m �� �أ�ÿ �K �� �ز������cȡ�ӷ����Pȥ����������10��20cm���xȡ������ �� �� ϵ �� �� �w�����ö��䷨�ɼ����H������ͬ һ�ӵظ�ȡ���c���������ϲ���̖̎����ͬ�r�ɼ��Ǹ��H�����M�И�̖̎�����������b����Ў���������(4 ��)����.�������r���H�������ھ���Y�x���Ǹ��H���Ӵ��L���^�Y���M���������|�y ��(��1)���� �� ��;� �� �t ���������������|��ȡ��Ҏ(gu��)�y����ʽ��ȫ �לy�������f�R����ɫ������Ч�לy�������}��-������ᷨ���ЙC�|�y���������t�� �-�� �� �� ����ȫ �� �y �����ð����P�Ϸ���ˮ��y �� �� �� �A ��-�U ɢ ����ȫ⛜y�����ÉA��-�����ȷ�����Ч⛜y�����������@����-�����ȷ���pH �y�������λ��[28].
����1.1.2 ���B(y��ng)��
���������[29]ָ�����������}����Դ�r������������@������������Դ��ͣ���˲����������}�o�C�����B(y��ng)��(������10.0g�������@0.5g�������V0.3g���Ȼ��c0.3g���Ȼ��0.3g�����၆�F0.03g�������i0.03g���������}5.0g����֬18.0g�����sˮ1L��pH7.0��7.5)�� �� �x �� �o �C �� �� ��.�����ɽ����ЙC�����B(y��ng)��(������10.0g�������@0.5g�������V0.3g���Ȼ��c0.3g���Ȼ��0.3g�����၆�F0.03g�������i0.03g������֬0.2g��̼���}5.0g����֬18.0g�����sˮ1L��pH7.0��7.5)�����x���ЙC����[30].�l(f��) �ͻ��A���B(y��ng)���x�� LBҺ�w���B(y��ng)��(�ȵ�����10.0g����ĸ������5.0g���Ȼ��c0.5g��pH7.0��7.2)[31].
����1.2 �����ĺY�x���b�������B(y��ng)�l����(y��u)��
����1.2.1 �������x�c����
����ȡ�u���� ʎ �� �� �� �� �� Һ �� �� Һ �� �� ��(10-3��10-4��10-5)ϡጣ�Ϳ���������}�o�C�����B(y��ng)�����ɽ����ЙC �� ƽ �� �� �B(y��ng) �� �ϣ�ÿ �� �� �� �O ��3�M �� �ͣ�����28 �������B(y��ng) �� ��(�� �� �C �� �� �� �B(y��ng)3d���� �o �C�����B(y��ng)7d)[32-33]��ӛ � �� �� �� Ȧ �� �� ��.�� �� �� �������Ä����������B(y��ng)3��7d�������ξ����D����ţ �� �� �� �� �� б �� �� �B(y��ng) �� �ϣ��� �� �� 4 �� �� �����[34].
����1.2.2 �����Y�x
���������Y�x����ƽ����Y�c�uƿ�ͺY.ƽ����Y��ӛ䛸����������Ȧֱ������ �� ֱ �� �� �� �� �� ֵ�������� �O5�M �� �ͣ�ȡ ƽ �� ֵ.�u ƿ �� �Y �� �� �� �Y �õ��ľ���ӷN��Һ�w���B(y��ng)���У��u�����B(y��ng)7d(28 �棬160r/min)���� �� �B(y��ng) �� �� �� �� �M �� �x �� ̎ �� (4 �棬10000r/min��10 min)�������f�R����ɫ���z�y����Һ�е����������Д���������������.
����1.2.3 16SrDNA �y���b��
��������16SrDNA ͨ���������� F27�� R1492���Y�x�õ��ļ����M�ДU���������������Ϻ��~������Ƽ�����˾�M �Мy�� �y �� �Y �� �� �� �� RDP(http����rdp.cme.msu.edu/)�� NCBI(http����www.ncbi.nlm.nih.gov/)�� �� �� �� �M �� �� �� �� �� �� �����xȡ�c GenBank��ͬԴ����ߵ����У������b������.
����1.2.4 ���B(y��ng)��(y��u)��
�������B(y��ng)���M�� ��(y��u) ������ �� �� ͬ ̼ Դ(�� �� �ǡ��� �ǡ����ǡ��� �� �� �� �ۡ��� ѿ �ǡ��� ¶ ��)�� �� �� �A �� �B(y��ng) ���е�̼Դ��������Һ��600nm �µ������(A600)�_����� ̼ Դ.�� ׃ �� �� ̼ Դ �| �� �� �� (0.5%��1.0%��1.5%)�M�о������B(y��ng)�Դ_����ѝ��.�� �� �� Դ(�����@���� �� �@���� �� ⛡��� �� �ˡ��� ĸ �ۡ��� ��)�� �� ���m�|���֔�(0.5%��0.8%��1.0%��1.3%��1.5%)�Ĵ_����ȡ��ͬ��ʽ.
�������B(y��ng)�l����(y��u) �����������l����׃����r���� �e ��׃���� �l(f��) �� Һ �� pH(5.0��6.0��6.5��7.0��7.2��7.5��8.0��9.0)���bҺ��(10��20��30��40��60mL����100mL�l(f��)��ƿ)���� �N �� (1%��3%��5%��7%��9%���� �� �w �e �֔�)�����B(y��ng)�ض�(20��25��28��30��35��40 ��)���y �� �� �� ��A600���_�����������mֵ.
��������ԇ�� �� �� �� �� �� � �Y �����O Ӌ �� �� �� ��ˮƽ����ԇ���ڳ�ʼpH ֵ(A)���bҺ��(B)���ӷN��(C)�����B(y��ng)�ض�(D)�Լ���ͬˮƽ�����B(y��ng)�l���M�Ѓ�(y��u)����ÿ��̎���O3���؏�.
����1.3 ����̎��
�������� Exce12010ܛ���M��ԭʼ�������������������D���L�ƣ��\��SPSS19.0ܛ���M�І����ط������(�@��ˮƽ0.05)�� �� С �@ �� �� ��(LSD)�� �� �� ���^������ Pearson���Pϵ�����_�� pH �c �� �� �� ֮ �g�����P�Pϵ���\ �� �� �� �O Ӌ �� �֢� v3.1̎ �� �� �� ԇ��.
����2 �Y���c����
����2.1 �����ĺY�x�Y��
���������x���ľ����M���ΑB(t��i)�����Д࣬������ʡ�֘I(y��)�ƌW�о�Ժ�ΑB(t��i)�W�b�����������B(y��ng)�ΑB(t��i)���l(f��)�F4�N��ͬ���g��ɼľ���H���������ΑB(t��i)����.���Y�x���Ľ����Բ����İ�ɫ�����ɫ���\�Sɫ����������ʈA�Ρ���Ҏ(gu��) �t �Σ�߅ �� �� �� �� �R���� �g �� �� �� �������� �� �� �� �� �� ��.��ͬ��������L�ٶȲ�һ�£��^�������������L�ٶ��^�죬����24h�����L��������;������ڽ��������48h����L���^�ã����L�ٶ��^����.
������ͬ���gɼľ���H�����������N�����ڲ(��2).�Ĕ���������ɼľ�˹����¸��H��o�C���Ĕ���������2.12×105��3.64×105cfu/g(cfu������γɆ�λ)�����ЙC���Ĕ���������1.31×105��2.14×105cfu/g.���У�15aɼľ���H�����Ľ�o�C�������ͽ��ЙC���������@�������������g(p<0.05)���� � �� �� �� �࣬�����ЙC����͔��� 9���c4aɼľ����͔���ͬ.
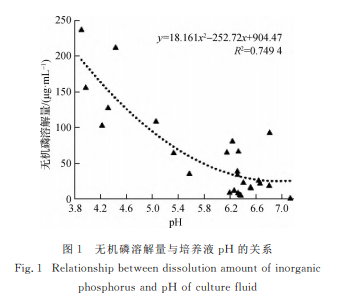
���������� �� �� �� �� �� �x �� �� Ч �� �� �� �@ �� ��(y��u) �� �ľ����M����������Ч�����M��16SrDNA �y���b ��.ͨ �^ �� ɼ ľ �� �H �� �� �� �M �� ƽ �� �� �Y���o �C�����B(y��ng)�����ЙC�����B(y��ng)���о����F�^���@����Ȧ���������H�� �� �� �� �� �� �� �� �o �C �� �� �� �C �� �� �D�����ɱ�ֲ��������������õ���Ч����.���о����Y�x�õ������^���@���õ�25���o�C�����20��� �� �C �� �� ��.�� �� �� �� �� �M �� �u ƿ �� �Y �� �l(f��) �F���ڹ��w���B(y��ng) �� �� �� �� Ȧ ֱ �� �c �� �� ֱ �� �� ֵ �� �� ���꣬��Һ�w���B(y��ng)���еĽ�����������һ���@��ͻ�����@�c�S�i�w��[35]�P�ڽ����ڹ��wƽ���Һ�w���B(y��ng)�ɷN��ʽ�� �� �� �� Ч �� �� �� �� �� �� �� �P ϵ �� �� �� �Y��һ��.�D�Д�����ĸ��ͬ��ʾ��@��(p<0.05)����ͬ.�D2 ��o�C�����������Fig.2 Phosphorussolubilizingcapacityofinorganicphosphorus-solubilizingstrain
����������ͨ�^�����ЙC����ܽ��y�����ף��l(f��)��Һ��pH �� �ͣ��� �� �� �� �� ӳ �� �� �� �� �� �� �� �� �ߵ�[36-37].���о� �� ̽ �� �� �o �C �� �� �� �� �� �� �� �� �r�����������B(y��ng)Һ��pH �M�Мy�������yֵ�����ڌ��սM���@һ�Y���Mһ����C�������YՓ.�l(f��)��Һ �ğo�C���ܽ����cpH �ʘO�@��ؓ���P��(p<0.01��R2=0.7497��r=-0.8659)�������������ľ������B(y��ng) Һ pH �^ �ͣ���֮pH �^�ߣ�ֻ�Ђ��e����ʬF�(�D1).�ЙC���ܽ��� �c pH �� �� �P �P ϵ �� �� �@ ��(p>0.05)���� �ЙC�������ҺpH ��Ҫ������6.58��7.42֮�g���H���� Y3��Һ��pH ��1.89���ʬF�������ԣ��c���������^����ԭ���������ڴֽ�����ø�����[38]���� Y3�Է����ЙC����l(f��)�]��������[39-40].
����ͨ�^�ͺY�����Æ����ط������������ğo�C���ܽ���.�o �C �� �� �� �� �^ �� �� �� �� �M(CK)�� �� �� ��(�D2)������������4.01��235.88μg/mL���� �� �� ������6.31��238.08μg/mL.W1�Ľ���Ч���@��������������(p<0.05)���������_238.08μg/mL;�� ��o�C��Ч�������� W15���������H��6.31μg/mL.W1�������� W15��������37.73��.
�����ЙC���ܽ����^�հ��սM��������(�D3)������������1.07��11.33μg/mL��������������4.78��15.04μg/mL.Y9�Ľ���Ч���@��������������(p<0.05)���� �� �� �_ 15.04μg/mL;����Ч��������Y10�������� �H ��4.78μg/mL.Y9�� �� �� �� Y10��������3.15��.
����2.2 �����b���Y��
������ PCR �U �� �� �� 16SrDNA �� �� �� �� (�� �(http����jxmu.xmu.edu.cn/upload/html/20220115.html)�DS1�� S2)ͨ �^ �� �� BLAST �� �� �� �� �õ����ƾ���ĵ��̖�c���ƶ�.���b����W1�鲻�ӗU����(Acinetobactersp.�����̖ MZ145066.1)(�D4)��Y9����ײ��Ͼ���(Klebsiellasp.�����̖ MZ145064.1)(�D5).
����2.3 �������B(y��ng)�l����(y��u)��
�����C���� �� �� � �Y �����x �� �� �o �C �� Ч �� �� �� ��W1�c���ЙC��Ч����õ� Y9�M�����B(y��ng)���M�փ�(y��u)�������B(y��ng)�l����(y��u)��.�D5 ���� Y9��ϵ�y(t��ng)�l(f��)����Fig.5 PhylogenetictreeofstrainY9
����2.3.1 �����،��Y�x
�����������L�����ڲ�̼ͬԴ���� Դ���� ʼ pH���b Һ�����ӷN�����ضȵ�푑����@���(�D6).̼Դ�������M����ꐴ��x�Ȼ�ӵ���Ҫ������Դ[41].6�N�� ͬ ̼ Դ(�� �� �ǡ��� �ǡ��� ѿ �ǡ��� �ǡ���¶���Ϳ��� �� �� ��)�� W1�� Y9�� �� �L �� �� Ӱ 푲�@����W1�� Y9�������Ǟ�̼Դ�r��A600ֵ���@����������̼Դ�r���������LЧ�����(�D6(a))����Ξ��¶������ѿ�ǡ����ǡ������Ե��ۡ�����.�ʸ�׃���m̼Դ�����ǵ��|���� ��(�D6(b))��W1��1.0%�r���L���@�����������|���֔��r���� Y9��0.5%�r���L�����.
������ԴҲ���������L�l(f��)������ҪԪ��֮һ.���Խ�ĸ�۞�Ψһ��Դ�r��W1�� Y9�����L���@������������Դ�r(�D6(c))���� �� �� �� �� �ˣ��� �� ĸ �� �� �� ���ˌ��������L����Ӱ��@����������4�N��Դ.���O�ú���ͬ�|���֔���ĸ�۵����B(y��ng)�����Y������ W1��1.50% �r���L���@����������̎������ Y9��1.25%�r���L���� ��(�D 6(d))���� �| �� �� �� �� �� �r �� �� ��A600ֵ׃��� �� �� �� �@���� �� ԓ �� �� �� �� �� �� �� ĸ�۵�Դ׃����������.
����pH8.0 ̎ �� �� W1 �� �L �� �� �� pH 7.0 ��7.5̎���² �� �� �� �@ �����ʺ��m(x��)����ԇ��ȡ����3�N��ʼpH ̎��;�� Y9���L����Ҫ��pH5.0��6.0֮�g���� �� څ �ݣ��� �m(x��) A600ֵ �S �� pH �� �� �� ����(�D6(e)).�� ͬ �b Һ �� �£��� �� �� �L �� �@ �� �� ͬ��W1�c Y9�����m�bҺ������30mL(�D6(f)).��ͬ�ӷN���£�3%��5%��7%�ӷN���� W1��ҺA600ֵ����@������ Y9��9%�� �N �� �r �� �L �� �@ �� �� �� �� �� ̎��.�ض��^���^��Ҳ�����ھ������ �L��35 ��̎ ���� W1���L���@����������̎������ Y9���L����28��30��35 ��̎���µIJ���@��������OӋ�� �� ԇ ��Mһ���_�����m�ض�.
����2.3.2 ����ԇ�
�����OӋ����ԇ�x��4������(��ʼ pH ֵ���bҺ�����ӷN���͜� ��)�� ÿ �� �� �� �O ��3�� ˮ ƽ.W1������ A ���ʼpH ֵ(7.0��7.5��8.0)������ B ���bҺ��(20��30��40mL)������ C ��ӷN��(3%��5%��7%)������ D �� �� ��(30��35��40 ��).�� �У��� �� A�� W1���L��Ӱ��������C�� D��֮��W1��������B(y��ng)�l����A2B2C1D3������ʼpH7.5���bҺ��30mL���ӷN��3%���ض�40 ��(��3).
����Y9������ A ���ʼpH ֵ(5.0��6.0��6.5)������ B���b Һ �� (20��30 �� 40 mL)���� �� C �� �� �N ��(5%��7%��9%)������ D ��ض�(28��30��35 ��).����RD>RB>RC >RA��4�����،��������L��Ӱ푳̶Ȟ�ض�>�bҺ��>�ӷN��>��ʼpH���t Y9������� �B(y��ng) �l �� �� A2B1C3D1���� �� ʼ pH ��6.0���b Һ ��20mL���ӷN��9%���ض�28 ��(��4).
��������ԇ�Y���c�����،��Y�����ڲ������������֮�g���ڽ������ã�������ԇ�Y�����߂�ʴ_��.
����3 ӑՓ�c�YՓ
�����澳��r�£�ֲ���ܭh(hu��n)��Ӱ푕����l(f��)�����đ�����.�u�@����[42]���ɼľ��ϵͨ�^���������푪�������{ �ȣ��ӿ�������L�ԑ������h(hu��n)��.�� �� �� �� Ч�����S���r����ľͨ�^�����{��(ji��)��ɫ@ȡ���ĠI�B(y��ng)Ԫ�أ���̎�ڵ����{�ȗl���µ���ľ�������đ������⣬߀��Ҫ����������Ƅ������ПoЧ��Ԫ�ص��D���Ա��C���o.
�����Ϸ��^(q��)ɼľ�˹�����������Ч���ڹ̶����õ�Ӱ��º������ͣ����H��������đ����܉���Ч���������[43].���о���ɼľ���H�Y�x���Ľ�o�C�� W1�� �� �� �� �� �_238.08μg/mL���� �� �C �� ��Y9���������_15.04μg/mL.Y9������ЙC���ܽ������ڷ���ȫ��[44]��ɼľ���H�Y�x���Ğ������ϲ��˻����¾�(Burkholderiaubonensis)�Ľ���P5(��������195.61mg/L)���@��������鲻ͬ�N�������x�C�ƾ��ж����ԣ��M�����·�����N͔����IJ�ͬ��Ӱ푽� �� �� �� �� �� �� �� ��[45];�� Ҳ �� �� ��ָ������������Ŀ��g���|��[46]�Լ�����h(hu��n)��[47]�IJ�����������cֲ�ﻥ���a��Ӱ�.
���������������������M�� Blast���ȵõ� W1�鲻�ӗU���٣�Y9����ײ��Ͼ���.���ĵ�[48]������ӗU���������r�I(y��)�Б�����������ص��ܽ����������Y�x����JL-1������������118.04mg/L������Ч���@������ W1.���ɵ�[49]�� �� �� �� �� �� �� �� �� ��(K. pneumoniae)�� �� �� �� ֲ �� �� �� �� �� �� �� �� �� Ҫ ���ã����о��ĽY����C�˸�Ч���� �����������@��.�f�贵�[46]���Y�x���IJ��ӗU���� PsbM8�� ��ؽӔM�Ͻ�(Arabidopsisthaliana)��ϵ���������@������Ȧ���F���Mһ���ó����ӗU�����ڽ�������ĺY�x�c�b���о��Б����^�V���ĽYՓ.Ŀǰ��Ҋ����Ľ�����Ҫ�й̵�����(Azotobacter)���نΰ�����(Pseudomonas)��ѿ �� �U �� ��(Bacillus)���W �� �Ͼ���(Erwinia)����������(Bradyrhizobium)����ù��(Penicillium)���� ù �� (Rhizopus)�� � ù �� ��(Streptomyces)[9��34��46].��һ�����ᘌ���ͬ�N�ٽ����Ľ���Ч����M��̽��.
�����x�ø�Ч�����M�����B(y��ng)���M�ֺ����B(y��ng)�l����(y��u)����������������m������h(hu��n) �������M��l(f��)�]���Ч�����պ����g���H��Ҏ(gu��)ģ���õ춨���A.���B(y��ng)����(y��u)�����Y �� �� ����W1 �� �� �c Y9 �� �� �� �e �� 1.0% ��0.5% �������Ǟ����̼Դ���@�c���ĵ�[50]����IJ��ӗU�������̼Դ�������ǵĽYՓһ��;��Դ������x��քe��1.50%��1.25%�Ľ�ĸ��.�Ϸ��t���^(q��)������ԣ�Y9����ƫ�����ԭh(hu��n)�����m�ˑ������Ϸ�ɼľ���H;�� W1����ƫ��A�ԭh(hu��n)�������ڌ��H�����^���м����{��(ji��)������A�ȣ��� �� �� �L ��(chu��ng) �� �� �m �h(hu��n) ��.�f �˻۵�[51]��ɼľ���H�Y�x���������ϲ��˻����¾���P5�������L��pH ���m������5��6�����m�ضȞ�25��30��.Ŀǰ�l(f��)�F�Ľ������(y��u)���B(y��ng)�l��ƫ��ߜحh(hu��n)������ �� �� �� �� Ч �� �� �c �� �� �� �� �� �P��� �� ɼ ľ ���a�^(q��)�����^�͵���r����Ч����Ŀǰ�F�Ѓ�(y��u)�ݽ������(y��u)���B(y��ng)�ض���ֵ��̽���ķ���.
�����S�������cֲ�ﻥ���C���о�����څ���ƣ����H������ �� �Q �� ľ �� �H �I �B(y��ng) Ԫ �� �{ �� �� �} �I �� �еđ���ǰ�������ӏV�.�F���о���Ҫ̽ӑ�����ĺY�x���b�������B(y��ng)�l����(y��u)���������ÙC�ƣ����ګ@�þ����� �� �� �L �����l(f��)�]��Ч���������Ч��.�� ־ ����[52]��������ЙC�|������̼���ȵ��ǽ�������l(f��)�]���õ���������.���˸��õ،�����������ڌ��H���a������һ���о��Б�����̽ӑ�����cֲ��Ļ� �� �C �ƣ���ͨ�^�m���{�حh(hu��n)�����ӣ��� �� �� �� ���� �� �� �桢���L�����ṩ������YԴ���������� �M��ӷN�����ķ�ֳ�c���L���Ķ�������������Ƶ����{�ȭh(hu��n)���ṩ�(y��u)�ľ��N�YԴ֧��.——Փ�����ߣ��w ��1��������1��������1���S ��1���dzе�2���� �I1*
���������īI��
����[1] �����£��μt.ֲ���ϵ푑������{�ȵęC���о�[J].�����ƌW��2015��27(3)��389-397.
����[2] �S�_������ٻ���� �� ������.һ��������ĺY�x���b �� �����������B(y��ng)�l���ă�(y��u)��[J].���\�g ͨ ��2015��31(2)��173-178.
����[3] �����־S���O������.�����^(q��)�ֵ�������Ч�������ķ��x���Y�x����� �� Ч ��[J].�|���֘I(y��)��W�W��2013��41(11)��83-85��122.
����[4] �߹��P���h�����̿£���.1��������b����Ӱ��������������[J].�|�� �� �I(y��) �� �W �W ��2020��48(1)��102-104��109.
����[5] ���G�t�������֣�܊����.�L��ʩ�ʼt����ˮ����������׃�������� �� ӯ ̝ �� � �� [J].�� �� �W ��2017��54(6)��1471-1485.
����[6] ղ���b��ꐷ�������С�w����.�Ё��Ꭷ����t���^(q��)ɭ����������A������������Ч��[J].���B(t��i) �W ��2009��29(9)��4673-4680.
����[7] ���܊�������������ϣ���.���������ɼľ�����~�|���pʧ���B(y��ng)�ֺ�����Ӱ�[J].�����c�h(hu��n)�� �� �� �W ��2018��24(6)��1211-1220.
����[8] ���粨����ʬ����dzе�����.�����{���������������ɼľ������ �� �� �� �� Ӱ � [J].�F �� �r �I(y��) �� �W��2020��48(1)��108-113.
����[9] �_�P������Ӱ�������ã���.�B��ɼľ���H����砵�������� Ⱥ �� ׃ �� Ҏ(gu��) �� [J].�� �B(t��i) �W �s ־��2020��39(9)��2921-2922.
����[10] �����������ʩ �l(w��i) ������.�˵��������������������������ΑB(t��i)�D�� �{ �� �� �� �� ��[J].�� ����2020��52(4)��668-675.
����[11] �S�Иˣ����^ᓣ��� �c �ˣ���.�dɼ���cɼľ�B���ֵ������������|�� �� ľ �� �L �� �� �^[J].�� �I(y��) �� �W��2010��46(4)��1-7.
����[12] ��Ę壬������Ԭ �^ ������.�����{���¼ӹ茦�������ڹ衢�נI�B(y��ng)���~�G�؟ɹⅢ����Ӱ� [J].ˮ �� �� �� �W��2017��31(1)��303-309.