�l���r�g�����ٷ���@�ֹ��̎��g�[��1��
ժ Ҫ�� ժ Ҫ: ��Ȼ�l���������y���~Ƭ��ϵ�y�� �⻯�WЧ�� ( Fv /Fm)���F���@��׃���� ���� Fv / Fm�S���յ����������� ,�� 14: 00�����_���ֵ������S���⏊�Ĝp�� Fv / Fm�����֏͡�һ�����~�S��ѭ�h�P�I�M�������S�| ( Z)�����c Fv / Fm��ؓ���P���ö����K�Ǵ� ( DTT)��
����ժ Ҫ: ��Ȼ�l���������y���~Ƭ��ϵ�y�� �⻯�WЧ�� ( Fv /Fm)���F���@��׃���� ���� Fv / Fm�S���յ����������� ,�� 14: 00�����_���ֵ������S���⏊�Ĝp�� Fv / Fm�����֏͡�һ�����~�S��ѭ�h�P�I�M�������S�| ( Z)�����c Fv / Fm��ؓ���P���ö����K�Ǵ� ( DTT)��� Z ���γɺ� ,�����Ƴ̶ȴ���� �Y�������c�~�S��ѭ�h���P�ķ�ݗ��������ɢ�������Ǯa�������Ƶ�ԭ��֮һ������̎��ǰ�ι D1���ϳ����Ƅ��ֿ�ù�� ( LM) , Fv /Fm�½��������� ,�����ڰ����c�����µĻ֏ͳ��F� ,������̎���������� LM,�t�� Fv /Fm �ɷN�֏͟o���@Ӱ푡� �@��������������y���~Ƭ D1���Ŀ������D��ֲ���w���@�N D1�����g�����ͻ�֮�g�ĄӑBƽ������DZ��⏊��p���ı��o�Է����� ��Ȼ�l�������{�Ȳ����܌����y���~Ƭ D1���ă��pʧ��
�����P�I�~: ������; PS�� �⻯�WЧ��;�~�S��ѭ�h ; D1����;�y��
���������{����ֲ�������õĹ������ܵ�Խ��Խ���о��ߵ��Pע�� �����ǽ�����S����Ȼɫ�ط�ݗ��������ɢ�^�̵İl�F ,�˂��������ƙC�����J�R�Ѱl���˺ܴ�ĸ�׃ (�S��ȫ 1992, Long& Humphries 1994)�� �҂����^�y�����������y���~Ƭ�Ї��ع�����ݬF�� ,���Ɯy�c���������P (�ϑc�� 1995) ,��һ���� Fv /Fm�½��̶Ⱥ��~�S��ѭ�h���M�ֺ������Pϵ ,�e�Ǻ������S�| ( Z)���PϵĿǰ߀��������P�� D1�������D ( turnov er) �c�����Ƶ��Pϵһֱ���f������ Ohad( 1984)�J���ϵ�y�� �������ĵ��g���c D1�������P�� D1�������D�l���ڹ������^�̵������A�� ,�� Clela nd( 1990)�t�J���ϵ�y�� ʧ��l���� D1���׃��pʧ֮ǰ , D1�������Ǟ�����ֹ��ϵ�y������ʧ��̶ȵ��Mһ��������B�� ( 1994, 1996)����Ȼ�l����ɺ���估���gС�����^�y��δ�l�F D1���ă��pʧ�����Č��о��y���~Ƭ������ù����ƵęC�� ,����̽ӑ�~�S��ѭ�h�� D1�������D�c�����Ƶ��Pϵ��
����1 �����c����
����1. 1 ��������
�����������y������Ȼ�l�����������ԡ���ȏ� 45 cm,��� 60 cm,�b���� 16. 5 kg ,��Ҏ������
����1. 2 �~�G�؟ɹⅢ���Ĝy��
�������ñ�yʽֲ��Ч�ʷ����x (Ӣ�� Ha nsa tech��˾���� )�y����ʼ�ɹ� ( Fo) ,���ɹ� ( Fm)��ϵ�y��⻯�WЧ�� ( Fv /Fm)�� �yǰ�A�� 5 min,�W��r�g 2 s��
����1. 3 �~�S��ѭ�h���M�ַ���
������ֱ�� 1 cm �Ĵ����ȡ�~�AƬ 8�� 10Ƭ ,�������� ,�����SʯӢɰ�c CaCO3 ,�� 100% ��ͪ�гɄ�{�� 1000 r /min�x�� 5 min�� �~�S��ѭ�h���M�ֵĸ߉�Һ��ɫ�V���������w���� ( 1995)������ �~�G�� a( chl a)���~�G�� b( chl b)������ UV -120�ֹ���Ӌ�y����
����1. 4 �~Ƭ���w���Q�����Ĝy��
����������� ( Pn)��������ͨ���ܶ� ( PFD)�ɱ�yʽ��Ϛ��w����ϵ�y (Ӣ�� ADC��˾���� )�@�á� �D�����Д����� 8�� 10Ƭ�~��ƽ��ֵ��
����1. 5 ���Ƅ�������
�������x���~Ƭ���~����ˮ���ټ�һ�� ,Ѹ���D���� 30 mmo l /L�Ķ����K�Ǵ� ( DT T)�� 3 mmo l /L���ֿ�ù�� ( LM)��Һ�� ,�քe������ (�s 30μmol· m - 2· s - 1 )�l���·��� 10 �� 3 h�� �����~Ƭ�������sˮ�С�
����2 �Y ��
����2. 1 �y���~Ƭ������ʼ��~�G�؟ɹⅢ����׃��
�����ĈD 1���Կ��� ,�y���~Ƭ������� ( Pn)��׃���������p���͡� Pn��һ�θ߷���F������ 10: 00���ҡ��˕r�⏊ ( PFD)�s�� 1200μmo l· m - 2 · s - 1 ������ 14: 00ǰ�� , Pn�_���ֵ (�⏊�s 1600μmo l· m - 2· s - 1 )���˺� Pn���л��� ,�� 16: 00�_�ڶ��θ߷� ,Ȼ���S���⏊�Ĝp�������͡�
������ϵ�y�� �⻯�WЧ�� ( Fv /Fm)�t�S���յ����������͡��ȳ��F�� 14: 00���� ,�c�糿 ( 6: 00)��� ,�½� 21. 8%�������� 18: 00�֏� 90%���ϡ����ɹ� FmҲ���F���Ƶ�׃��څ�ݡ��c���෴ , Fo������ ,�� ,��׃�����Ȳ��� (�D 2)�� Fv /Fm���½���Ҫ�� Fm ��������
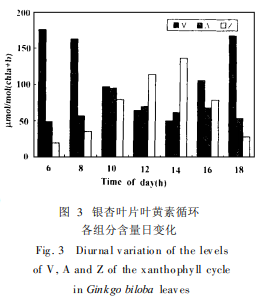
����2. 2 �~�S��ѭ�h���M�ֺ�����׃��
�����D 3��ʾ�~�S��ѭ�h���M�����S�| ( V)���h�������S�| ( A)�������S�| ( Z)������׃���������S�� PFD���� ,�����Ƽ��� (�D 2)�����S�| ( V )��Ó�h������Ҳ�S֮�ӏ� , Z�ĺ������ߡ����� Fv /Fm������͕r , Z�ĺ���Ҳ�_���ֵ�� ����S���⏊�p�� , Z�ĺ����½������ϽY������ ,�c�~�S��ѭ�h���P�ķ�ݗ��������ɢ�����ӿ����ǹ�ϵ�y��⻯�WЧ���½���ԭ��֮һ
����2. 3 �~�S��ѭ�h���Ƅ� D TT���y���~Ƭ�����Ƶ�Ӱ�
���������K�Ǵ� ( DT T)�����~Ƭ�� ,�c�����~Ƭͬ�r���ڏ��� ( 1600 μmol· m - 2· s - 1 ) ��̎�� 2 h ,�ٜy���~Ƭ�ğɹⅢ�����֏��^�� ,�Y����D 4���c������� , DTT̎�����~Ƭ Fv /Fm�½����������� 29% �� �������������S�|���γɺ�����Ƴ̶ȼ��ء� ���� DTT ̎�����~Ƭ ,�ڰ��������»֏� ( 50 μmo l· m - 2· s - 1 )�̶Ȳ�ͬ�� �y���~Ƭ�ڰ�̎�IJ���ȫ�֏� (�֏ͼs 67% )���������~�S��ѭ�h�� ,�������� D1���׃����⡣
����2. 4 D1���ϳ����Ƅ� LM ���~Ƭ�����Ƽ���֏͵�Ӱ�
�����D 4 DTT̎�팦�y���~Ƭ�����Ƴ̶ȼ��֏͵�Ӱ� Fig . 4 Pho toinhibitio n a nd its r eco ve ries under DTT trea tment ���Mһ��̽ӑ���⌦�y���~Ƭ D1����Ӱ� ,�҂��^���� D1���ϳ����Ƅ��ֿ�ù�� ( LM )���~Ƭ�����Ƽ���֏��^�̵�Ӱ푡� �l�F�ι LM ���~Ƭ���� 2 h ���� ( 1600 μmo l· m - 2· s - 1 )����� , Fv /Fm���½��̶ȱȌ��������� 39. 7% ,���Ұ��з��� 16 h �� ,ֻ�֏��� 25% (�D 5)�� �����˕r��������� D1�����Ɖġ� ���ڻ֏��A������ LM ���֏ͼ����֏;����_ 95%���ϡ� �f�� 2 h����̎����δ�l�� D1���ă��pʧ��
����3 ӑ Փ
������ 50��� Hag er�l�F��������ľGɫֲ���~Ƭ�����~�S�غ�����׃���ԁ� ,�~�S��ѭ�h��������ٵı��o�����ѵõ������о��ߵ��Jͬ ,���H���S�|��Ó�h�����ܺ�ɢ�^ʣ���l�� ,��������� ( Demmig-Adamas 1990, 1992) , Z����Ҳ���c�~�G�wɫ�؏ͺ����������ɢ�� Z�ĺ����c�ǹ⻯�W⧜�ʾ������P ( Lo ng a nd Humphries)����һ�����C����������^�쵽�y���~Ƭ������M���������S�|�����c Fv /Fm�½������������P (�D 3) , �ι DT T�� Fv /Fm�½��̶����� (�D 4) ,�����Ƽ��� ,������ D1���ēpʧ ,������ه�~�S��ѭ�h�ķ�ݗ��������ɢ���������y���~Ƭ�a�������Ƶ�ԭ��֮һ��
����D1�������Dһֱ�ǹ����ƙC���о������^���S���I�� ,������ϡ��������h���l���IJ�ͬ ,�����˂���δ�_�ɹ��R�� �҂�ͨ�^���^����̎��ǰ������ D1���ϳ����� ( LM )�����ƻ֏��^�̵IJ ,�l�F����̎��ǰ���� LM����� D1���דpʧ (���֏Ͳ���ȫ ) ,������̎�������� LM,���֏ͼ�����֏͟o���@� (�D 5) ,�����μ��ď����{��δ���� D1���ă��pʧ�� �ɴ˿�֪ ,�����{���´��� D1�������D ,����Ȼ�l���� D1�����g�����ͻ��_���ӑBƽ�Ⲣ������ D1���ă����⡣ �������Ϸ��� ,�҂��Ɯy��Ȼ�l�����y���~Ƭ�~�S��ѭ�h����ķ�ݗ��������ɢ�� D1���Ŀ���ʧ���nj��¹�ϵ�y��⻯�WЧ���½��ăɂ���Ҫ���ء�——Փ�����ߣ��� �� ,�ϑc�� ,�w���� ,�S�L�� ,�u ��
���������īI:
����[1 ] �S��ȫ ,������,���s�.ֲ�������õĹ����� [ J].ֲ������WͨӍ , 1992, 28( 4): 237�� 243.
����[2 ] LON G S P, HUM PHRIES S. Pho toinhibiti on of phot osynthesis in nature [ J]. Annu. Rev. Plant. Physiol. Plant Mol. Biol. , 1994, 45: 633�� 662.
����[3 ] �ϑc�� ,����ϼ,�w���ܵ� .�y�ӹ�����Ե��о� [ J].�֘I�ƌW , 1995, 31( 1): 69�� 71.
����[4 ] O HAD I, KYLE D J, ARN TZEA C J. M emb rane protein damage and repair, removal and replacement of inactivat ed 32-kil odalton polypeptides. in chlo ropl ast membranes[ J]. J. Cell. Biol. , 1984, 99: 481�� 485.
����[5 ] CLEL AND R E, DEM MIG ADAMS B, ADAMS W W �� et al. Phos ph orylati on stat e of th e ligh tharvesting chl oroph yll prot ein complex of ph ot os yst em�� and chl orophyll fluorescence charact eristics in Monstera d eliciosa Liebm. and Glycine ma x ( L. ) [ J]. Merrill in res ponse to ligh t. Aust. J. Plant Physiol. , 1990, 17: 589�� 599.
����[6 ] ���B�� ,�S��ȫ.��Ȼ�l����ɺ�����~Ƭ������õĹ����� [ J].ֲ������W�� , 1994, 20( 1): 46�� 54.
����[7 ] ���B�� ,�S��ȫ,�����.���gС���~Ƭ������õĹ����Ʋ����S D1���ă����� [ J].ֲ��W�� , 1996, 38( 3): 196�� 202.
����[8 ] �w���� ,�ϑc��,�S�L�ǵ� .ֲ��M�����~�S��ѭ�h�M�ֵĸ�ЧҺ��ɫ�V������ [ J].ֲ������WͨӍ , 1995, 31 ( 6): 438�� 442.
����[9 ] DEM MIG ADAM S B. Carot enoid and phot oprot ection in pl an ts: A role f or the Xanth oph yll Zeaxanthin [ J]. Biochimica et Biophysica Acta. , 1990, 1020: 1�� 24.
����[10 ] DEM MIG ADAMS B, ADAM S W W�� . Phot oprot ection and oth er res ponses of plants to hi gh li ght stress [ J]. Annu. Rev. Plant Physiol. Plant Mol. Biol. , 1992, 43: 59�� 62