�l���r�g��2020-03-05���ٷ���t�WՓ���g�[��1��
ժ Ҫ�� ժҪ: �������������ɶ�������������ͨ�����ӽM�ɵġ����{�ؼ������ݻ�����_��һ����أ�ԓ�^�����ܶȵ��D����ӡ��o���Ӽ����������P���^����������������ӵĮ����D䛻��S���[�������������P��Ҫ���[������ͨ�^�M�b�������������ӣ��@����
����ժҪ: �������������ɶ�������������ͨ�����ӽM�ɵġ����{�ؼ������ݻ�����_��һ����أ�ԓ�^�����ܶȵ��D����ӡ��o���Ӽ����������P���^����������������ӵĮ����D䛻��S���[�������������P��Ҫ���[������ͨ�^�M�b�������������ӣ��@�����M��N��������_���Ķ������[����������ֳ�����u���D�Ƶ�����;���Ƴ��������ӵĻ��ԣ��t�@�������[�����������L�ʹ����Č�Ŀǰ������[�������г��������ӵĽY�����������{�أ��Լ�����������ˎ���аl�F���M���˿��Y��ּ�ڞ��аl�µ�ᘌ����������Ӟ���c�Ŀ��[��ˎ���ṩ��Փ���A�ͽ��b��
�����P�I�~: ������;����������;�D�;���Y
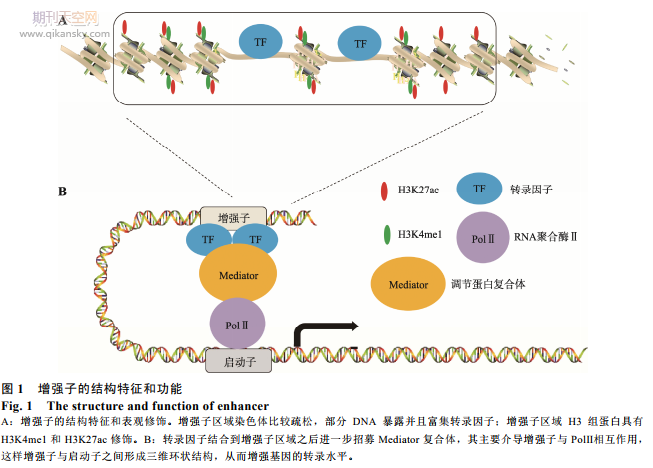
����20 ���o 80 ������о��l�F SV40 ������һ�� DNA ���Ќ��ڼ���(Oryctolagus cuniculus)�� β-�鵰��(β-globin��һ�N�܉�ͨ�^�F߲���h�����ԽY�����ĺ����Ե����|)���D䛾����������ã���ˌ��@һ�� DNA �Q��������(enhancer)[1]���S����о��l�F�ڲ�����:����Ҳ����������Ե� DNA ���У������h���x���o�����Ե����������D�[2~4]���� 30 ���о��C�������Ӿ�����������(�D 1)[2~5]��(1) ������ DNA ����̎��Ⱦɫ�w���ɵą^���c��С�w�нM��������D����ӵĽY�����P;(2) �����ӻ����c�� DNA ���нY�ϵĽM���� H3 �ĵ� 4 λه����μ���(H3K4me1)�͵� 27 λه����������(H3K27ac) ��̶ȳ������P[6];(3) �����Ӱl�]������Ҫ�����Ӆ^��͆����ӵą^���ֱ������ã��γ����S�h��Y��(3D-loop)�������Ӻ͆����ӵ�������ɶ�N���錧���� Mediator �ͺ��w��Cohesin ��[6,7]��
�������]��x���[������������Ͷ����Щ�W�g��ڿ�
�����S�� DNA �y���g�İlչ��������������Mһ�����J�R�����������ӵ��о�ҲԽ��Խ���롣2013 �꣬���� Young R.A.����(Whitehead Institute for Biomedical Research)���ڮ��r�����ӵ��о��״��������������(super enhancers��SEs)�@һ��������l�F��̥�ɼ���(embryonic stem cells��ESC)����Ҫ�D����ӽY����һЩ������������ϣ��@Щ����������ӌ��ھS����̥�ɼ����ĸ������P��Ҫ�������@Щ����������Ӷ��x�鳬��������[8]�����������Ӻ����f�����ɶ��������ӽM�ɵ�һ����أ��������ܶȵ��D����ӡ��o���Ӻ������ӱ��^���������ͨ�����������д�С���D����ӵĽY���ܶȡ������D䛵������Լ����D��������Ƅ��������Ծ���ͬ[8]���S����о����H�l�F���������Ӵ����ڶ�N��������У�Ҳ�Mһ�����_�˳��������Ӆ^�e����ͨ�����ӵĹ�������(�D 2)[6]��(1) ���������Ӿ��и��ܶȵ� H3K27ac �� H3K4me1 ����Լ� Mediator �ͺ��w�� Bromodomain containing 4 ����(BRD4���c�M�������������λ�c�Y��)�ĽY��;(2) ���������ӽY�ϵ��D������Լ��c�D䛻������P��Ⱦɫ�w�Ę�ӛ����ͨ�����Ӹߺܶ�;(3) �����������{�صĻ������ͨ�������{�صĻ�����_ˮƽ�ߺܶ�;(4) �M�ɳ��������ӵĆ�������Ҳ��������ͨ������һ�Ӽ�������D�;(5) ���������ӿ��ԽY�ϽM�����خ����D�����;(6) �c��ͨ��������ȣ����������ӻ��Ԍ����D����ӵ���������[9,10]���@Щ�F��֧��һ�����f�����������Ӱl�]������Ҫ�Y�ϵ������������ϵ��D����ӵĺ����fͬ�����д����D����ӽY�ϵ������ӌ��ڻ����D䛵��{�ؕ����D����ӝ�ȵĸ�׃������[11](�D 2)����Ȥ���ǣ������ڳ����������ϵ���Ҫ���D�����Ҳ�ܳ��������ӵ��{���D䛣��@����ζ�������������{�ػ����D䛴����������fͬ���ã�Ҳ���γ��˼����еĺ����D��{�حh· (core transcription regulatory circuitry, CRC)[12,13]���������ڳ����������{�ػ�����_�����Ժ��������ԣ�������܉�f�{���������L���l�����ֻ��ͼ����ȸ��N��B���^��[9,14~16]��������Ҫ�ij����������c�[���������Pϵ�����[�������е��{���Լ�ԓ���cˎ�����[���ί��еĬF���@ 3 �������U���������������[�������е����á�
����1 �����������c�[�����Pϵ
����2013 �꣬Young R.A.���ڰl�F�ڶ�l�Թ����������г��������ӵą^��ļ���˸ߝ�ȵ� Mediator �ͺ��w�� BRD4[17]���@��ζ���ڶ�l�Թ����������г���������̎�ڻ��B�����ܷ����������������������{�صĻ���(MYC��IRF4��PRDM1��XBP1) ���ڶ�l�Թ������İl���Ͱlչ���P�I�Ĵ��M����[17]�����m�о��l�F�������������ڶ�N�[���о��Ј�����珛���Դ� B �����ܰ���[18]��T ���������ܰͼ�����Ѫ��[19,20]��Ĭ�ˠ�������[21]����������Ѫ��[22]��С�����ΰ�[10]���ѳ���[23]����Ƥ��[24]���[�����[25]����ɫ����[15]�����ٰ�[26]��ʳ���[�����[27]�ͽY�c��[28]�ȡ����[�������г����������{�ص��P�I�������������������Dz����_�ģ��@����ʾ����������ͨ�^�{���@Щ��������[�����ɺ��[�����ԾS�����P�I����[9,29,30]�����ڼ����ڰ�׃�^���д�����ij����������������γɾ��й����Ե�Ԫ������˳��������ӵĻ�������鼚����׃��һ�N��־[9,29,30]���C������Ŀǰ�о����������������ӵļ�����Դ�ʹ�����������[�������Đ����D�������������Ӳ����������a��������D䛼������ã����Ǿ��a������ microRNA (miRNA��һ�N�L�ȼs 22nt ��С RNA)���D䛼�����Ҳ�����{�ع��ܡ�������ʡ�����WԺ����ϵ Phillip A. S.�����о��M���� CRISPR/Cas9 ����M�������l�F���������Ӳ��H���M miRNA ���D䛣�Ҳ����ͨ�^��ļ Drosha/DGCR8 ���ͺ��w���Mǰ�w miRNA (primiRNA)�ij��죬�Դˁ��{�ؼ����N��خ��� miRNA ������[31]���� 18 �N�[�����������l�F������Щ�[�������г��������ӻ������{������Щ�[�������г��������ӻ����½����Mһ�����������ڼ�����׃�^���м���ij��������������c�ٰ� miRNA ���P����ʧ��ij�����������Ҫ�{���ְ� miRNA ������[31]�������о���ʾ���{�� miRNA �ij��������ӻ����c�[���l���lչ�������P����ˣ������������϶��� miRNA (SE-miRNA)���Н����ɞ鼚����׃�������־��[31]�������[���������\���Լ��ί�������Ҫ���R�����x������֮�⣬����������߀�����{���L朷Ǿ��a RNA (long non-coding RNA, lncRNA)���D�[32]�����[������M���аl�F�ܳ����������{�ص� lncRNA LINC01503 ���@���{���Mһ���о��l�F�� LINC01503 �ı��_ˮƽ�c�[����������A���ؓ���P��LINC01503 �߱��_�IJ��������ʵ͡��о���������������ӻ������Ӆ^��Ҳ���Ա��D䛮a�� RNA���Q�� enhancer RNA (eRNA)��eRNA ���ԅfͬ���������Ӽ����D�[33,34]�����������Ӱl�]���ܲ��H��ه�ں͆�����֮�g�� 3D-loop ���γɣ�Ҳ��ه�ڳ����������D䛵� eRNA �����ɡ���ˣ����R���Ͽ��ԽY�� lncRNA �Լ� eRNA ��ˮƽ�������M�о����ί���
�����[���Į��|�Ժܴ�һ����������һ���[���ȵļ���ͨ�����܁�Դ�ڶ�����ͬ�ļ�����¡�����@Щ��ͬ��¡��Դ���[�������䳬�������ӵļ���Ҳ���ڲ���@�͞�^���[�����ͻ��[��������Ⱥ�ṩ��һ�N�µ��b�����������磬ͨ�^�����ķ��������ܼ����������ﻯ�W���z���W��������ֳ� 4 �����͡�����ͨ�^���@ 4 �����͵������ӈD�V�����l�F��һ�N�µā��ͣ��@�N���͵ij��ܼ����������ж������c�[�����|�����P�ij���������Ⱥ[35]��������Ҫ����ͨ�^�������@��[�������ij����������{�ص��D����ӿ������_�����خ��Եĺ����D��{�حh·(CRC)��ͨ�^���� CRC �����_���� LIM homeobox transcription factor 1 alpha (LMX1A��һ�N�D�����)�ڵ� 4 ��͵ij��ܼ�������һ����Ҫ�D�����(master transcription factor)[36]��ͬ�ӣ������������|����Ҳ�l�F�����r������������ٰ����������������{�ص��خ��ԵĻ���Ⱥ���S�ּ������L����ֳ[26]����Ҋͨ�^���ڲ�ͬ�[�������������ӵĈD�V�������Ԫ����A�y�[�����ͣ��l�F֮ǰ�ί��IJ����Լ��µĝ����ί����c�����[���ί��ṩ��˼·���·���[6]��
�����C���������ڶ�N�[�������о��l�F����������̎�ڮ��������B���䌦�ڰл�����{�سʶ��ӻ������M mRNA �����ɡ����M miRNA ���D��Լ����졢���M lncRNA ���D������Լ����������������D����ɵ� eRNA ���������Ҳ�fͬ���á�����֮�⣬ͨ�^�L���[�������������ӈD�V�����A�y�[�����ͣ�������|���[���ṩ�yһ���ί�ƽ�_��
����2 �[�������г��������ӵ��{��
�������[�������г��������ӵ��{������Ό��F����?���ڌ���С����̥�ɼ����l�����о����һ��ģ�ͣ��M�ɳ��������ӵ�ÿһ�������Ӷ����л��ԣ������������ӵĹ��������һ��ƽ�_���@��ƽ�_�R�����c�l�����P����̖ͨ·���f�^������̖���@Щ��̖�fͬ�{�س��������ӻ��Ԇ��ӻ����D�(�D 3)[11]��ͬ�ӣ��c���������P�ij���������Ҳ�������[��������ه����̖ͨ·���D����ӡ��� Wnt ��̖ͨ·��������ĽY�c�������У����P�ij��������Ӆ^���˺ܶ��� Wnt ��̖ͨ·�K�˵��D����� 4 (transcription factor 4, TCF4)��ͨ�^����������� Wnt ��̖ͨ·�����Կ��Ƴ����������{�صĻ����D�[11,37]���ڴƼ������w(estrogen receptor, ER)��Ե����ٰ������У����P�ij��������Ӆ^��ۼ��˴����� ERα;������������ٰ�������ȱ��̴����صı��_���c�����P�ij��������Ӆ^������ȫ��ͬ���D�����[26,37]��
�������[�������У���̖ͨ·�Ķ�挦���������ӵĻ����M���{�ء�2015 ������������W Licht J.D. ���ڈF��о��l�F Ras-Erk �����c���������ӵĻ����������P������ Ras ���Ļ��ԕ����³������������P������(�� H3K27ac)��ʧ�������½����������P�����D�;���� Ras ���������{�ذ�����ij��������ӻ���[38]����һ���棬�ٰ���̖ͨ·����ͨ�^�ٿv�D䛙C���{�����������ӵĻ��ԡ��D䛕�ͣ�Ǽ���� RNA �ۺ�ø��(RNA polymerase ��Pol ��) �چ����Ӹ���ֹͣ�D䛵�һ�N��B[39]���������ĸμ����У�Hippo ��̖ͨ·����ͨ�^���ƕ�ͣ�� Pol �� ጷţ���������������ӻ��������{�صĻ����D�[40]��Ȼ�����ڸΰ������� Hippo ��̖ͨ·��ȱʧ���� YAP(Yes associated protein)��ˣ�YAP ���Y�ϵ������������ϣ���ļ Mediator �ͺ��w�ͼ�����������ه�Լ�ø 9(Cyclin-dependent kinase 9�� CDK9)��ʹ��ͣ�� Pol ���M�뵽�����B�����M�������D�[41]����ˣ��ڸΰ��� YAP ͨ�^����������Ӵ��M��������D䛡�
���������о��������������ӿ��������B�Ӱ�������̖ͨ·�;S���[���������ԵĻ����D䛱��_��������Ȼ���Mһ���о��l�F��̖ͨ·���ڳ��������ӵ��{���c�D������ڳ��������Ӆ^�ӑB�Y�����P�����磬�� NOTCH1 �������µ� T ������Ѫ��(T-ALL) �����У�NOTCH1 �ڻ���M�Ͼ����ձ�ĽY�ϣ�����ֻ�в��� 10%�Ļ����� NOTCH1 ��̖ͨ·�ĸ�׃�Б��𣬶��@Щ�������� NOTCH1 �Y���ڌ����ij����������ϡ�����@Щλ�c�Gʧ NOTCH1 �ĽY�Ͼ͕����³��������ӵ�������ʧ[42]��
SCISSCIAHCI